RS/tRNA Foundational Publication Support
Amiram, Miriam, Adrian D. Haimovich, Chenguang Fan, Yane-Shih Wang, Hans-Rudolf Aerni, Ioanna Ntai, Daniel W. Moonan, et al. (dec) 2015. “Evolution Of Translation Machinery In Recoded Bacteria Enables Multi-Site Incorporation Of Nonstandard Amino Acids”. Nature Biotechnology 33: 1272-1279. doi:10.1038/nbt.3372.
Wang, Lei, Zhiwen Zhang, Ansgar Brock, and Peter G Schultz. (2003) 2003. “Addition Of The Keto Functional Group To The Genetic Code Of Escherichia Coli.”. Proceedings Of The National Academy Of Sciences Of The United States Of America 100 (1): 56-61.
Young, Travis S, Insha Ahmad, Jun A Yin, and Peter G Schultz. (2010) 2010. “An Enhanced System For Unnatural Amino Acid Mutagenesis In E. Coli.”. Journal Of Molecular Biology 395 (2): 361-74. doi:10.1016/j.jmb.2009.10.030.
Young, Douglas D, Travis S Young, Michael Jahnz, Insha Ahmad, Glen Spraggon, and Peter G Schultz. (2011) 2011. “An Evolved Aminoacyl-Trna Synthetase With Atypical Polysubstrate Specificity.”. Biochemistry 50 (11): 1894-900. doi:10.1021/bi101929e.
RS/tRNA Usage Publications
Castro, Ana, Juan Manuel Carreño, James Duehr, Florian Krammer, and Ravi S. Kane. (aug) 2021. “Refocusing The Immune Response To Selected Epitopes On A Zika Virus Protein Antigen By Nanopatterning”. Advanced Healthcare Materials 10: e2002140. doi:10.1002/adhm.202002140.
Shapiro, Daniel Mark, Gunasheil Mandava, Sibel Ebru Yalcin, Pol Arranz-Gibert, Peter J. Dahl, Catharine Shipps, Yangqi Gu, et al. (feb) 2022. “Protein Nanowires With Tunable Functionality And Programmable Self-Assembly Using Sequence-Controlled Synthesis”. Nature Communications 13: 829. doi:10.1038/s41467-022-28206-x.
Stadler, Karina A., Walter Becker, Barbara Darnhofer, Ruth Birner-Gruenberger, and Klaus Zangger. (jul) 2022. “Overexpression Of Recombinant Proteins Containing Non-Canonical Amino Acids In Vibrio Natriegens: P-Azido-L-Phenylalanine As Coupling Site For 19F-Tags”. Amino Acids 54: 1041-1053. doi:10.1007/s00726-022-03148-2.
Li, Allyson, Rashmi Voleti, Minhee Lee, Dejan Gagoski, and Neel H. Shah. (mar) 2023. “High-Throughput Profiling Of Sequence Recognition By Tyrosine Kinases And Sh2 Domains Using Bacterial Peptide Display”. Elife 12: e82345. doi:10.7554/eLife.82345.
Bard, Jared, Charlene Bashore, Ken Dong, and Andreas Martin. (2019) 2019. “The 26S Proteasome Utilizes A Kinetic Gateway To Prioritize Substrate Degradation.”. Cell 177 (2): 286-298.e15. doi:10.1016/j.cell.2019.02.031.
Bard, Jared, and Andreas Martin. (2018) 2018. “Recombinant Expression, Unnatural Amino Acid Incorporation, And Site-Specific Labeling Of 26S Proteasomal Subcomplexes.”. Methods In Molecular Biology (Clifton, N.j.) 1844: 219-236. doi:10.1007/978-1-4939-8706-1_15.
Casilli, Fabrizio, Miquel Canyelles-Niño, Gerard Roelfes, and Lur Alonso-Cotchico. (2024) 2024. “Computation-Guided Engineering Of Distal Mutations In An Artificial Enzyme.”. Faraday Discussions 252: 262-278. doi:10.1039/d4fd00069b.
Ali, Bashir. (2024) 2024. “The N1 Domain Of The Peroxisomal Aaa-Atpase Pex6 Is Required For Pex15 Binding And Proper Assembly With Pex1.”. The Journal Of Biological Chemistry 300 (1): 105504. doi:10.1016/j.jbc.2023.105504.
RS/tRNA Pair Development Year
2015
ncAA(s) Incorporated
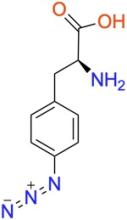
p-azido-L-phenylalanine (pAzF)
ncAA Structure (png, jpg, jpeg)
ncAA Utility
Used as a photocrosslinker, allowing for crosslinking and bioorthogonal click-chemistry ligation of proteins via strain-promoted cycloadditions (SPAAC) with suitably functionalized molecules.
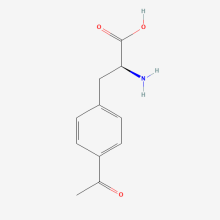
p-acetyl-L-phenylalanine
ncAA Structure (png, jpg, jpeg)
ncAA Utility
Site-directed spin labeling, electron paramagnetic resonance
RS Organism of Origin
Parent RS
RS Mutations
Y32L
D158G
E107T
F108Y
Q109M
I159C
L162R
R257G
D158G
E107T
F108Y
Q109M
I159C
L162R
R257G
tRNA Organism of Origin
Parent tRNA
tRNA Anticodon
CUA
Other tRNA Mutations
C17A
U17aG
U20C
G37A
U47G
U17aG
U20C
G37A
U47G
Multiple tRNAs?
N/A
RS/tRNA Availability
AddGene: Plasmid #73546
RS/tRNA Additional Notes
This RS/tRNA pair was evolved in the C321.ΔA E. coli strain from the AcetylPhe RS (pAcF-RS) originally reported in in 2002 foundational paper, used as a model RS in the 2010 foundational paper and characterized for permissivity in the 2011 foundational paper. In 2015 further residues were allowed to evolve to optimize p-azido-Phe and tRNA interactions and improve expression of multiple ncAA containing proteins using low RS levels. This evolved RS showed 25 fold higher expression of pAzF in GFP-3sites compared to the progenitor enzyme (pAcFRS) using a chromosome incorporated RS. Figure 4a of foundational paper shows that when included as multicopy plasmid it still highly outperforms the starting RS for incorporating 30 ncAAs in a construct, but that only outperforms the original unevolved RS by <2-fold when expressing GFP with 3 ncAAs, and that along with the improved expression it has quite a poor fidelity (about the same level of protein production in the absence of ncAA). Note that the "pAzRS" in this figure is the pCNFRS shown in the 2011 foundational paper to work very well for pAzF. The pAzFRS.2.t1 RS was also shown to be highly promiscuous, incorporating Phe variants pAcetyl, pIodo, pBromo, pChloro, pMethyl, ptrifluoromethyl along with methylTyr, 2-napthylAla and styrylAla.