RS/tRNA Foundational Publication Support
Chin, Jason W, Ashton Cropp, Christopher Anderson, Mridul Mukherji, Zhiwen Zhang, and Peter G Schultz. (2003) 2003. “An Expanded Eukaryotic Genetic Code.”. Science (New York, N.y.) 301 (5635): 964-7.
Takimoto, Jeffrey K, Katrina Adams, Zheng Xiang, and Lei Wang. (2009) 2009. “Improving Orthogonal Trna-Synthetase Recognition For Efficient Unnatural Amino Acid Incorporation And Application In Mammalian Cells.”. Molecular Biosystems 5 (9): 931-4. doi:10.1039/b904228h.
Chatterjee, Abhishek, Han Xiao, Michael Bollong, Hui-Wang Ai, and Peter G Schultz. (2013) 2013. “Efficient Viral Delivery System For Unnatural Amino Acid Mutagenesis In Mammalian Cells.”. Proceedings Of The National Academy Of Sciences Of The United States Of America 110 (29): 11803-8. doi:10.1073/pnas.1309584110.
Augustin, George, Fatema Bhinderwala, Nathan D Alexander, Iker Hernández, Stanislau Stanisheuski, Christina M Monnie, Alex J Eddins, et al. (2026) 2026. “Endogenous Site-Specific Encoding Of Trifluoromethyl-Bearing Phenylalanine And Tryptophan For In-Cell F Nmr.”. Journal Of The American Chemical Society 148 (7): 6744-6749. doi:10.1021/jacs.5c18349.
RS/tRNA Usage Publications
Anderson, Christopher, Ning Wu, Stephen W Santoro, Vishva Lakshman, David S King, and Peter G Schultz. (2004) 2004. “An Expanded Genetic Code With A Functional Quadruplet Codon.”. Proceedings Of The National Academy Of Sciences Of The United States Of America 101 (20): 7566-71.
Xiao, Han, Abhishek Chatterjee, Sei-hyun Choi, Krishna M Bajjuri, Subhash C Sinha, and Peter G Schultz. (2013) 2013. “Genetic Incorporation Of Multiple Unnatural Amino Acids Into Proteins In Mammalian Cells.”. Angewandte Chemie (International Ed. In English) 52 (52): 14080-3. doi:10.1002/anie.201308137.
Zheng, Yunan, Tommy L Lewis, Peter Igo, Franck Polleux, and Abhishek Chatterjee. (2017) 2017. “Virus-Enabled Optimization And Delivery Of The Genetic Machinery For Efficient Unnatural Amino Acid Mutagenesis In Mammalian Cells And Tissues.”. Acs Synthetic Biology 6 (1): 13-18. doi:10.1021/acssynbio.6b00092.
Italia, James S, Christopher Latour, Chester J J Wrobel, and Abhishek Chatterjee. (2018) 2018. “Resurrecting The Bacterial Tyrosyl-Trna Synthetase/Trna Pair For Expanding The Genetic Code Of Both E. Coli And Eukaryotes.”. Cell Chemical Biology 25 (10): 1304-1312.e5. doi:10.1016/j.chembiol.2018.07.002.
Osgood, Arianna O, Yunan Zheng, Soumya Jyoti Singha Roy, Nikolaos Biris, Myer Hussain, Conor Loynd, Delilah Jewel, James S Italia, and Abhishek Chatterjee. (2023) 2023. “An Efficient Opal-Suppressor Tryptophanyl Pair Creates New Routes For Simultaneously Incorporating Up To Three Distinct Noncanonical Amino Acids Into Proteins In Mammalian Cells.”. Angewandte Chemie (International Ed. In English) 62 (19): e202219269. doi:10.1002/anie.202219269.
Grasso, Katherine T, Soumya Jyoti Singha Roy, Arianna O Osgood, Megan Jin Rae Yeo, Chintan Soni, Christen M Hillenbrand, Elise D Ficaretta, and Abhishek Chatterjee. (2022) 2022. “A Facile Platform To Engineer Tyrosyl-Trna Synthetase Adds New Chemistries To The Eukaryotic Genetic Code, Including A Phosphotyrosine Mimic.”. Acs Central Science 8 (4): 483-492. doi:10.1021/acscentsci.1c01465.
RS/tRNA Pair Development Year
2003
ncAA(s) Incorporated
O-methyl-L-tyrosine
ncAA Structure (png, jpg, jpeg)
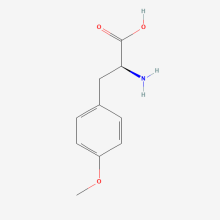
ncAA Utility
Labelling of muscles
(global)
(global)
m-Acetyl-L-phenylalanine
ncAA Structure (png, jpg, jpeg)
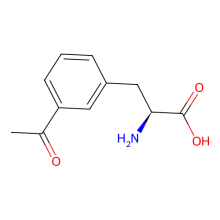
ncAA Utility
Labelling with
hydrazides, both in
vitro and on cell
surfaces
hydrazides, both in
vitro and on cell
surfaces
p-azido-L-phenylalanine (pAzF)
ncAA Structure (png, jpg, jpeg)
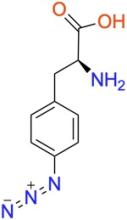
ncAA Utility
Used as a photocrosslinker, allowing for crosslinking and also for bioorthogonal click-chemistry ligation of proteins via strain-promoted cycloadditions (SPAAC) with suitably functionalized molecules.
For crosslinking, short wave UV light (~280 nm) is typically used. Upon absorbing a photon, the activated intermediate can crosslink to neighboring atoms, if present. If no crosslinking is possible, it will relax to a ground state that cannot be reactivated.
For crosslinking, short wave UV light (~280 nm) is typically used. Upon absorbing a photon, the activated intermediate can crosslink to neighboring atoms, if present. If no crosslinking is possible, it will relax to a ground state that cannot be reactivated.
p-iodo-L-phenylalanine
ncAA Structure (png, jpg, jpeg)
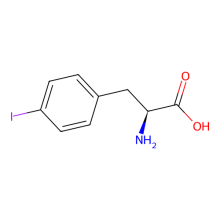
ncAA Utility
Can be used for phasing in protein crystallography.
p-bromo-L-phenylalanine
ncAA Structure (png, jpg, jpeg)
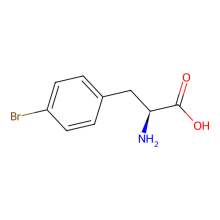
ncAA Utility
Mutant GFP
excitation/ emission
excitation/ emission
para-nitro-L-phenylalanine
ncAA Structure (png, jpg, jpeg)

ncAA Utility
n/a
2-Cyano-L-phenylalanine
ncAA Structure (png, jpg, jpeg)
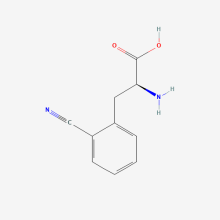
ncAA Utility
Potentially useful as fluorescence-based environmental sensor to study protein conformation. Also, can be an IR probe.
O-allyl-L-tyrosine
ncAA Structure (png, jpg, jpeg)
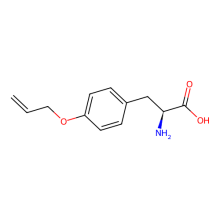
ncAA Utility
useful for thiol-ene click chemsitry
O-propargyl-L-tyrosine
ncAA Structure (png, jpg, jpeg)
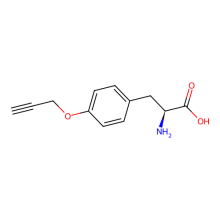
ncAA Utility
Reactive handle for azide-alkyne cycloaddition
trifluoromethyl-L phenylalanine
ncAA Structure (png, jpg, jpeg)
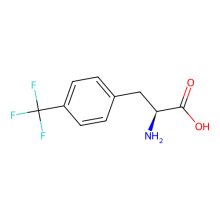
ncAA Utility
19F Probe for protein
NMR
NMR
RS Organism of Origin
Parent RS
RS Mutations
Y37V
D182S
F183M
D265R
D182S
F183M
D265R
tRNA Organism of Origin
Parent tRNA
tRNA Anticodon
CUA
Multiple tRNAs?
also works with Bacillus stearothermophilus Tyr tRNA(CUA)
RS/tRNA Availability
Available in Addgene Plasmid #50831, which also contains tRNA from both Bacillus stearothermophilus and E.coli.
RS/tRNA Additional Notes
Compared with original 2003 selected RS, D265R mutation was added based on 2009 foundational paper showing it improves recognition of the CUA anticodon. The 2013 foundational paper showed the RS to be highly permissive. With 1 mM O-me-Tyr in HEK293 cells, gave a purified yield of ~36 ug per 10^6 cells of eGFP(39), and was shown capable of incorporating 8 other ncAAs with yields ranging from 5 - 35 ug per 10^6 cells. It was also shown to function in multiple cell lines including primary cultures of fibroblasts and neuronal cells.
2026 foundational paper showed also functions well with p-trifluoromethyl-Phe. Incorporation into GFP(150) was ~80% efficient compared with WT expression, and at ~50% the level of pAzF incorporation. Incorporated into mulitple sites of cyclophilin A for in-cell NMR studies.
2026 foundational paper showed also functions well with p-trifluoromethyl-Phe. Incorporation into GFP(150) was ~80% efficient compared with WT expression, and at ~50% the level of pAzF incorporation. Incorporated into mulitple sites of cyclophilin A for in-cell NMR studies.